| |
| |
| |
| |
 父亲肺鳞癌,cT3N0M1a,ⅣA,脑转移
父亲2025年6月中旬因为头痛,一侧肢体不灵活入院,做了增强核磁,发现右侧额叶占位(3
父亲肺鳞癌,cT3N0M1a,ⅣA,脑转移
父亲2025年6月中旬因为头痛,一侧肢体不灵活入院,做了增强核磁,发现右侧额叶占位(3
 老公刚刚54岁,确诊肺癌,手术中发现
各位朋友好。
度过了慌乱的一个月,有机会发现了这个论坛,如获至宝,说说我家
老公刚刚54岁,确诊肺癌,手术中发现
各位朋友好。
度过了慌乱的一个月,有机会发现了这个论坛,如获至宝,说说我家
 SMARCA4-NSCLC 盲试靶向药有效!!
首先感慨一下今天真的是个好日子。努力了进三个月的治疗终于有了重大进展。
复查结果
SMARCA4-NSCLC 盲试靶向药有效!!
首先感慨一下今天真的是个好日子。努力了进三个月的治疗终于有了重大进展。
复查结果
 希望给没有阅读过的人一些体会
本文来自于此书,只是做一些摘抄,分享给深陷迷茫的人。文中观点真实与否,效果怎么
希望给没有阅读过的人一些体会
本文来自于此书,只是做一些摘抄,分享给深陷迷茫的人。文中观点真实与否,效果怎么
 L858R+S768I+TP53阿法替尼用药半年希
2023.11.23我父亲(63岁)因胸腔积液,呼吸困难,后背疼,声音沙哑入院。此前一个多月
L858R+S768I+TP53阿法替尼用药半年希
2023.11.23我父亲(63岁)因胸腔积液,呼吸困难,后背疼,声音沙哑入院。此前一个多月


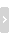


 显身卡
显身卡


